Obesity and type 2 diabetes are prevalent, chronic, metabolic conditions which have a significant impact upon the individual as well as on healthcare systems and economies. In addition to the daily burden of these conditions, obesity and type 2 diabetes increase the risk of developing serious long-term complications, such as heart attacks, kidney failure and lower limb amputations. The progressive nature of these conditions has driven the need to develop new, more efficacious medications, which allow patients to live healthier lives for longer.
Agonists of the glucagon-like peptide-1 receptor (GLP1R), such as semaglutide (Ozempic/Wegovy), have revolutionised the world of obesity and diabetes treatment. This is not only because they cause significant weight loss and improvement in glucose control, but also because dose escalation is permitted without the risk of hypoglycaemia.
More recently, tirzepatide, a dual agonist of the GLP1R and the glucose-dependent insulinotropic polypeptide receptor (GIPR), has shown even better results in people with obesity1 and type 2 diabetes.2 This dual agonism is curious, since GIPR agonism alone is associated with weight gain in rodents, and loss-of-function human variants in the GIPR are associated with lower body mass index.3

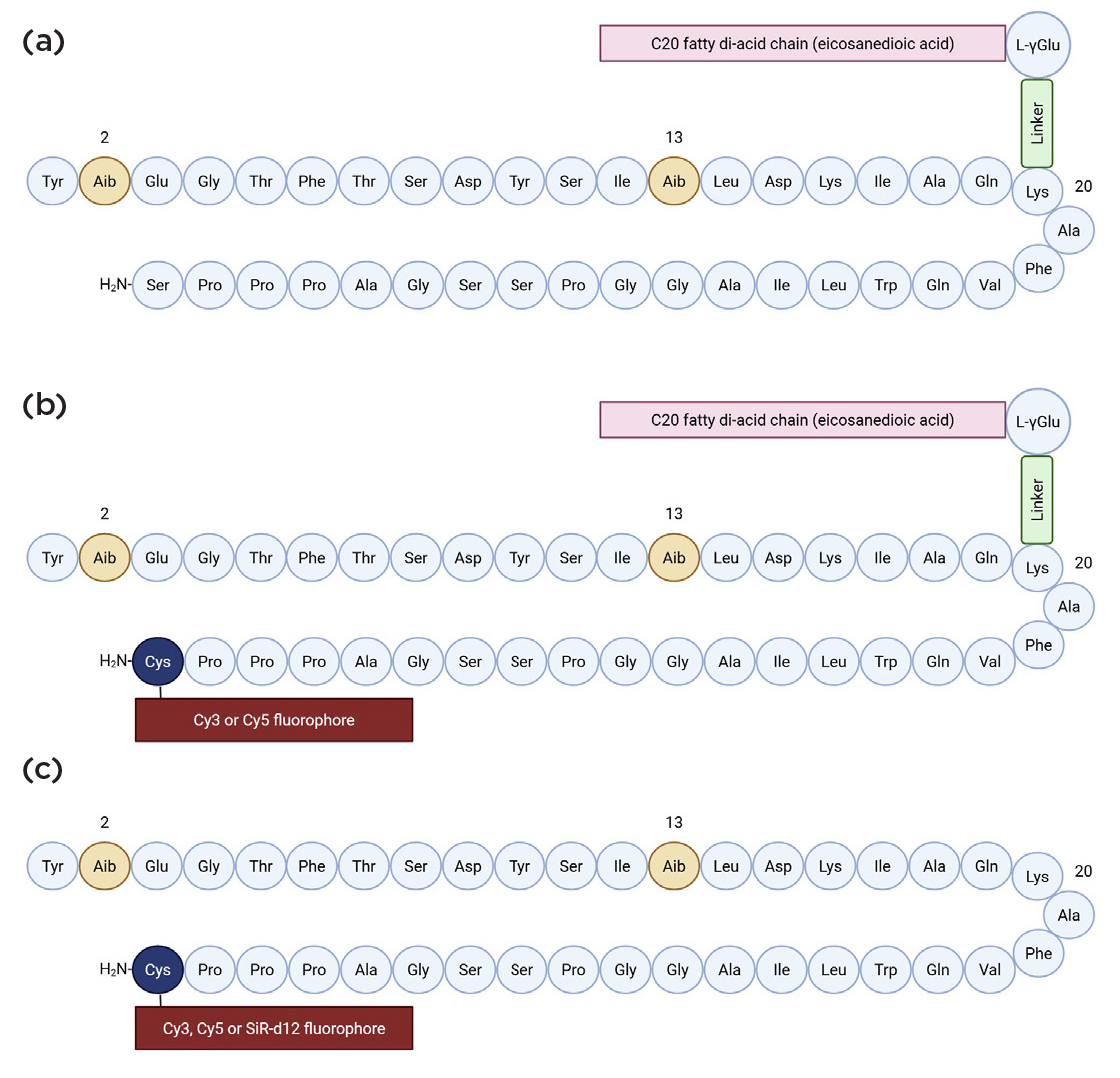
Figure 1. The chemical structures of (a) tirzepatide, (b) acylated daLUXendins544+/660+ and (c) non-acylated daLUXendin544/660/651-d12. The daLUXendin molecule is a mutant of tirzepatide, with a substitution of serine for cysteine at position 39 to allow for fluorophore conjugation, which can only be achieved with a C-terminal reactive amino acid such as cysteine. Images created using BioRender.com
QUESTIONS REGARDING DUAL GLP1R/GIPR AGONISM
Whilst the pharmacology of tirzepatide has been explored, there remains uncertainty regarding tirzepatide’s targets within the body, and whether differences in brain access play a role in its superior efficacy compared with GLP1R agonists.
There are many techniques to identify the presence of a receptor or its gene expression, such as reporter animals and antibodies. However, discordance has been reported between GLP1R gene expression and final protein expression.4 Furthermore, the GLP1R and GIPR are low-abundance receptors, and generating antibodies with sufficient sensitivity and specificity is challenging.5 Additionally, these techniques are unable to provide information on the brain and peripheral sites accessed by dual agonists, which is the key next step towards understanding their efficacy as well as informing future drug development.
daLUXendins TO EXPLORE DUAL GLP1R/GIPR AGONISM
To allow us to address these questions, we generated novel, fluorescent, dual GLP1R/GIPR probes, termed dual agonist LUXendins (daLUXendins), by coupling a fluorophore directly onto a modified tirzepatide molecule. We had previous success in using this coupling technique to generate a range of highly fluorescent and specific GLP1R probes, LUXendins.6 The coupling technique is versatile and permits the addition of fluorophores with a range of properties and colours to suit various experiments.
Acylated (+) and non-acylated red (daLUXendin544+/daLUXendin544) and far-red (daLUXendin660+/daLUXendin660) probes were synthesised for conventional imaging, as well as a probe adapted for super-resolution imaging (daLUX651-d12) (Figure 1).

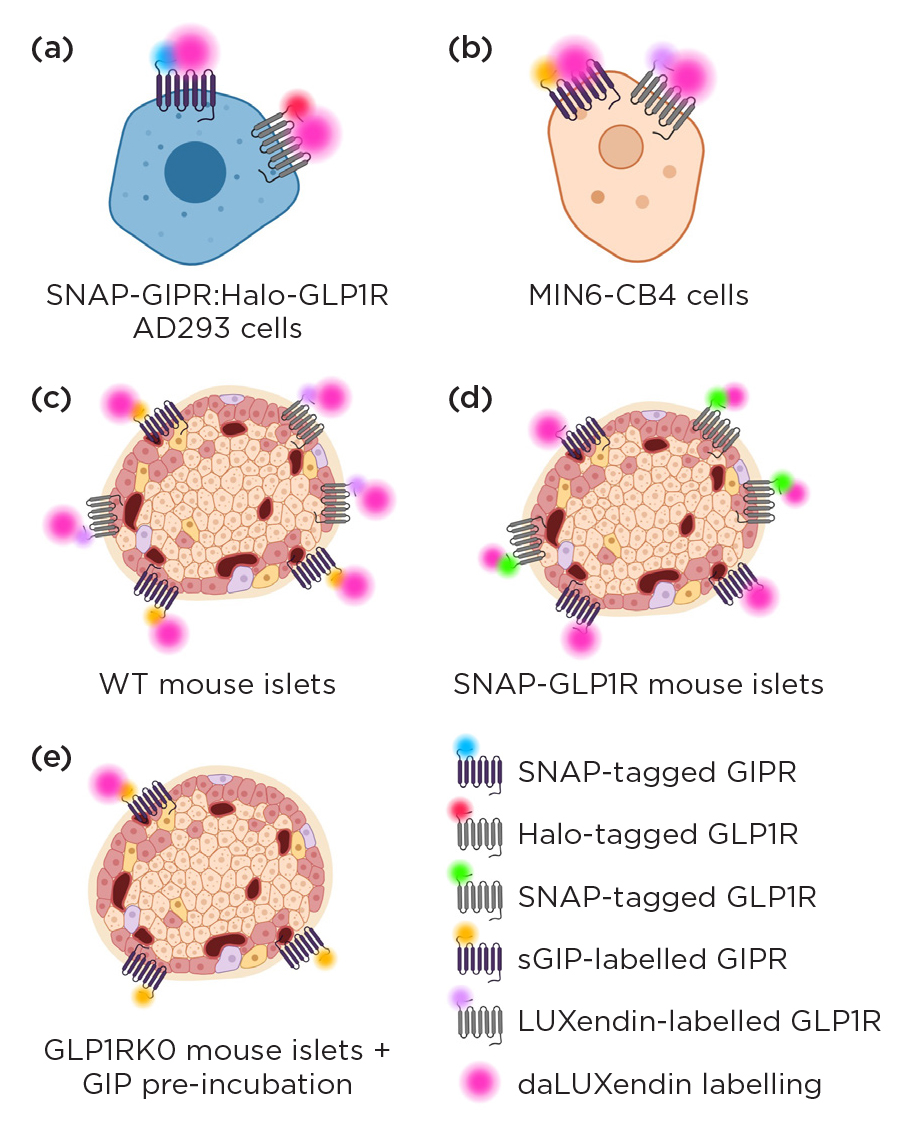
Figure 2. daLUXendins able to label GIPR and GLP1R when overexpressed & endogenously expressed. daLUXendin labels (pink) co-localised with labels for GIPR/SNAP-GIPR & GLP1R/Halo-GLP1R in (a) AD293 cells, (b) MIN6-CB4 cells (c) wild-type (WT) mouse islets. (d) daLUXendin labelled endogenously expressed GIPR & co-localised with SNAP-GLP1R in islets from the transgenic SNAP-GLP1R mice. (e) daLUXendin labels reduced by around 50% in islets from GLP1R ko mice & further by pre-incubation with excess GIP agonist.
TESTING OUR HYPOTHESES
We hypothesised that the superior efficacy of dual GLP1R/GIPR agonism over single GLP1R agonism could be due to:
• a different profile of receptor(s) engaged on synergistic cell and neurone types
• increased access into the brain, to more potently target appetite regulation centres
• differences in higher-order signalling events, such as preferential interactions between GLP1R and GIPR at the cell surface.
To interrogate each of these lines of investigation, we first assessed the daLUXendin probes pharmacologically and then validated their specificity in a range of relevant settings, including cell lines which over-expressed self-label enzyme-tagged GLP1R/GIPR, and mouse islets (Figure 2).
Both daLUXendin probes are potent GLP1R/GIPR agonists at mouse and human receptors. Tirzepatide is an imbalanced agonist, favouring GIPR over GLP1R in humans, but the opposite in mice.7 The chemical modifications made to daLUXendin increased functional selectivity at the mouse GIPR, allowing daLUXendin660 to confer more human-like properties on dual agonism in mice.
Differing from single GLP1R agonists, which primarily target β cells, daLUXendin labels α, β and δ cells in mouse islets and islets derived from human induced pluripotent stem cells. Further testing in mouse islets revealed that daLUXendin probes generate the formation of GLP1R/GIPR nanodomains, which are important for GLP1R signal amplification.8
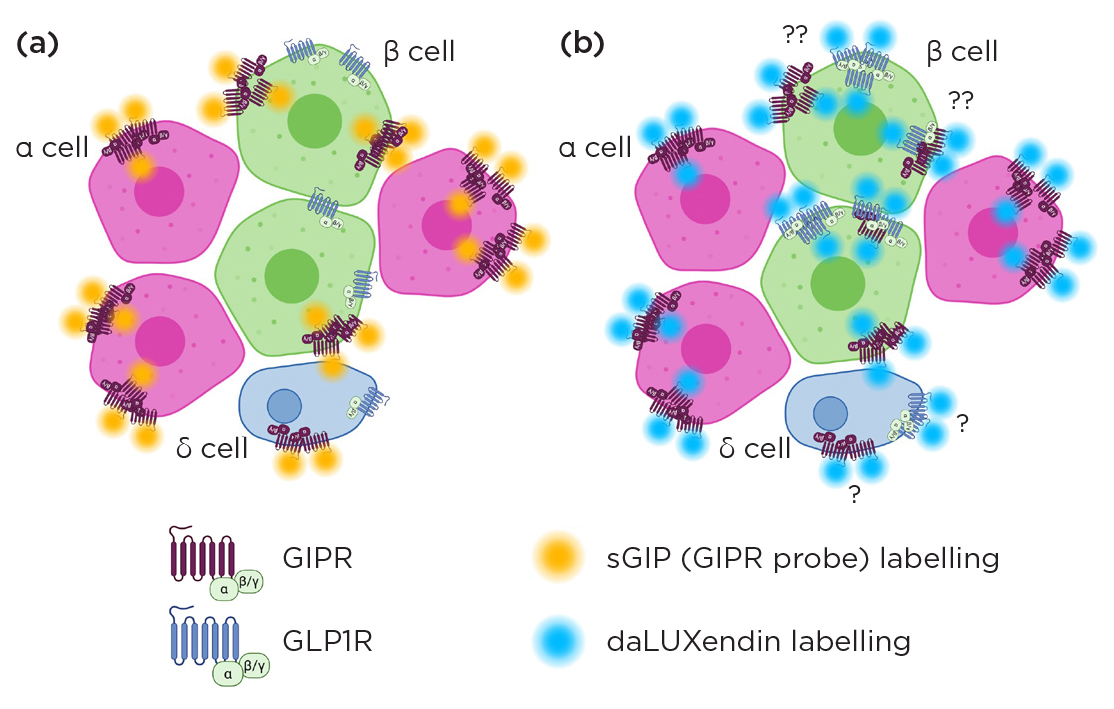
The number of nanodomains is increased compared to that seen with single GIPR agonist probes, but without change to the number of receptors engaged per nanodomain or the size of the nanodomain (Figure 3).
After peripheral administration, the depth of daLUXendin access to the brain was similar to that previously reported for single agonists and limited to the circumventricular organs, specialised areas of the brain with a reduced and plastic blood-brain barrier. Following intraventricular administration, daLUXendin probes were able to label tanycytes, specialised ependymal cells which line certain circumventricular organs, the third ventricle and hypothalamus, and which mediate the transport of peptides across these areas.

Figure 3. dSTORM imaging found (a) GIPR agonist probe sGIP engaged receptors into nanodomains - typically many more than the 3 receptors depicted in this simple schematic. (b) daLUXendin engaged more nanodomains. However, the number of receptors per nanodomain and size of nanodomains were the same. The presence of GLP1R on α and δ cells is unclear, as is whether the rise in nanodomain formation with daLUXendin is due to increased homotypic (GIPR–GIPR and GLP1R–GLP1R) or heterotypic (GIPR–GLP1R) interactions
Together, these results suggest that the superior efficacy of dual GLP1R/GIPR agonists stems from differences in the types of engagement and manner by which they engage islet cells, neurones and receptors, rather than the extent of brain penetration. In the islet, dual agonists engage α, β and δ cells, and therefore probably activate multiple paracrine mechanisms to enhance insulin secretion.9 Tanycytes probably convey dual agonists from the bloodstream to the cerebrospinal fluid to important feeding centres within the brain. Engagement of GLP1R/GIPR nanodomains is likely to contribute to signalling robustness within target organs.
THE NEXT STEPS
Going forward, functional interrogation of the islet cells, neurones and supporting cells targeted by dual GLP1R/GIPR agonists would help confirm their contribution to glucose lowering and food intake.
Additionally, agonists are under development that target the glucagon receptor (GCGR) in the form of triple GLP1R/GIPR/GCGR agonists, showing superior efficacy to dual GLP1R/GIPR agonists. The daLUXendin probes could be modified to incorporate GCGR action and address current uncertainties, such as how superior weight loss is achieved when GCGRs are considered to be largely absent in the brain.
ANNE DE BRAY
Academic Clinical Lecturer and Specialist Registrar in Diabetes and Endocrinology, OCDEM, University of Oxford, and Department of Metabolism and Systems Science, College of Medicine and Health, University of Birmingham
DAVID J HODSON
Robert Turner Professor of Diabetic Medicine, Director of The Bukhman Centre for Research Excellence in Type 1 Diabetes, OCDEM, University of Oxford
REFERENCES
1. Rodriguez PJ et al. 2024 JAMA Internal Medicine https://doi.org/10.1001/jamainternmed.2024.2525.
2. Frías JP et al. 2021 New England Journal of Medicine https://doi.org/10.1056/NEJMoa2107519.
3. Rosenkilde MM et al. 2025 Diabetes https://doi.org/10.2337/dbi24-0027.
4. Gray SM et al. 2020 Journal of Biological Chemistry https://doi.org/10.1074/jbc.RA120.014368.
5. Ast J et al. 2021 EBioMedicine https://doi.org/10.1016/j.ebiom.2021.103739.
6. Ast J et al. 2020 Nature Communications https://doi.org/10.1038/s41467-020-14309-w.
7. Willard FS et al. 2020 JCI Insight https://doi.org/10.1172/jci.insight.140532.
8. Buenaventura T et al. 2019 PLoS Biology https://doi.org/10.1371/journal.pbio.3000097.
9. Hartig SM & Cox AR 2020 Journal of Molecular Medicine https://doi.org/10.1007/s00109-020-01887-x.
All figures created in Biorender.com



{kind=link}
{kind=link}
{kind=link}