Karla Jade Suchacki gave the 2022 Early Career Prize Lecture for basic science at SfE BES 2022 in Harrogate. We are delighted to include this summary of her talk here.
Obesity and its associated co-morbidities, including cardiometabolic complications (e.g. type 2 diabetes mellitus, hypertension and cardiovascular disease), substantially contribute to negative global health outcomes. The key feature of obesity is increased white adipose tissue (WAT) mass; the role of ‘lesser known’ adipose depots, such as brown adipose tissue (BAT) and bone marrow adipose tissue (BMAT), is not clear in adult humans.
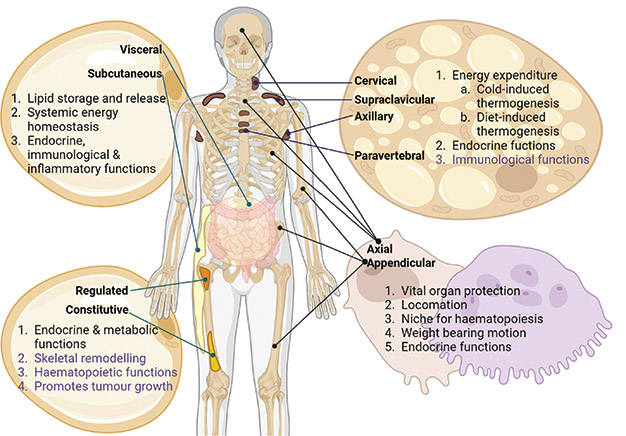
WAT is typically classified into visceral and subcutaneous depots that are dispersed throughout the body. As the primary organ of lipid storage, WAT is characterised by large monolocular lipid droplets, and plays a key role in systemic energy homoeostasis. It also has important endocrine and immunological roles (Figure).1

Figure. The distribution and function of WAT, BAT, BMAT and the skeleton. Each depot has specific roles as indicated, with putative functions highlighted in purple. In the skeleton, the bone-forming osteoblasts (pink) and bone-resorbing osteoclasts (purple) are shown. Created in BioRender
BROWN ADIPOSE TISSUE
In contrast to white adipocytes (WAds), brown adipocytes (BAds) contain multilocular lipid droplets, are rich in mitochondria and express uncoupling protein 1, which generates heat by uncoupling the electron transport chain. Therefore, BAT is a thermogenic organ that increases energy expenditure to maintain body temperature in a cold environment (Figure).
In adult humans, functional BAT was identified about 20 years ago, when positron emission tomography/computed tomography (PET/CT) using the metabolic tracer 18F-fluorodeoxyglucose (18F-FDG) led to its incidental identification.2 BAT is now known to be located in the neck, supraclavicular, paravertebral, peri-aortic and perirenal regions.
BAT activation improves insulin sensitivity and lipid clearance, highlighting its beneficial role in metabolic health. Furthermore, BAT activity is substantially reduced in obesity and type 2 diabetes mellitus. However, repeated cold exposure increases BAT mass and activity in subjects with obesity or type 2 diabetes mellitus, and can reduce WAT mass.3 Thus, activating BAT is an exciting approach to treating obesity and metabolic disease.
UNDERSTANDING BAT
Our understanding of human BAT remains in its infancy, and the regulation of human and murine BAT activation is not always preserved between the two species.4 Using transcriptomics of human BAds, we have recently identified the serotonergic system as a key regulator of human BAT, and inhibition of peripheral serotonin synthesis may be a novel strategy to treat obesity-associated metabolic disease.
'Inhibition of peripheral serotonin synthesis may be a novel strategy to treat obesity-associated metabolic disease.'
Furthermore, accurately quantifying BAT mass in humans is challenging, as estimates of BAT activity are based on substrate consumption, with no specific imaging biomarker for BAT reported. To address this, we have recently identified a novel PET tracer that can identify BAT without cold or drug stimulation in humans. In addition, novel PET tracers have the potential to provide mechanistic insights and improve our understanding of human BAT activation. To date, factors/agents that improve murine BAT function have failed to translate to humans, hence understanding of the regulation of human BAT is key to identifying novel pathways amenable to therapeutic manipulation.
BONE MARROW ADIPOSE TISSUE
BMAT is another unique adipose tissue depot that has been largely ignored since its identification over a century ago. Situated within the marrow cavity, BMAT constitutes approximately 70% of bone marrow volume (and approximately 10% of total adipose tissue mass) in healthy adult humans, principally in the appendicular skeleton.5
In contrast to WAds, bone marrow adipocytes (BMAds) proliferate in conditions such as anorexia nervosa and type 1 diabetes. BMAds exist in two subtypes: ‘constitutive’ BMAds that predominate at distal skeletal sites, and ‘regulated’ BMAds which are interspersed within the haematopoietic bone marrow at proximal skeletal sites and within the axial skeleton. BMAds are morphologically similar to WAds, having a large unilocular lipid droplet; they produce adipokines (leptin and adiponectin) and release free fatty acids in response to lipolytic stimuli (Figure).
THE ROLE OF BMAT
We recently showed that BMAT plays a key role in glucose clearance and is functionally distinct from WAT and BAT.6 Using 18F-FDG PET/CT, we revealed that BMAT resists both insulin- and cold-stimulated glucose uptake. Highlighting its key metabolic role, glucose uptake by BMAT is greater than is found in the axial bones or in WAT, and can even exceed skeletal muscle uptake.6
Finally, BMAT is situated within the bone marrow cavity and, through total body PET, we have recently determined that different bones within the skeleton have a unique glucose metabolism and form a complex metabolic network.7 In addition to the classical functions of the skeleton, both rodent and human studies have identified the skeleton as a key endocrine regulator of metabolism.8
These data highlight the need to broaden our focus beyond individual adipose depots, taking a whole body approach. As we continue to develop new analytical approaches to understand complex tissue interactions at a systems level, this will improve our ability to better understand mechanisms underlying metabolic disease and develop novel therapeutics.
Karla Jade Suchacki
Postdoctoral Research Fellow, Cardiovascular Science, Queen’s Medical Research Institute, University of Edinburgh
REFERENCES
1. Suchacki KJ et al. 2016 Current Opinion in Pharmacology 28 50–56.
2. Cohade C et al. 2003 Journal of Nuclear Medicine 44 1267–1270.
3. McNeill BT et al. 2021 European Journal of Endocrinology 184 R243–R259.
4. Ramage LE et al. 2016 Cell Metabolism 24 130–141.
5. Pachón-Peña G & Bredella MA 2022 Trends in Endocrinology & Metabolism 33 401–408.
6. Suchacki KJ et al. 2020 Nature Communications 11 3097.
7. Suchacki KJ et al. 2021 Frontiers in Medicine 8 740615.
8. Motyl KJ et al. 2017 Toxicologic Pathology 45 887–893.
Find out about all of the Society for Endocrinology's prizes and awards




{kind=link}