Follicle-stimulating hormone (FSH) is a glycoprotein hormone that plays a central role in reproduction by regulating ovarian follicle granulosa cells and testicular Sertoli cells which express the FSH receptor (FSHR). Anterior pituitary glycoprotein hormones include FSH, luteinising hormone (LH) and thyroid-stimulating hormone (TSH). Primates and equids express a placental glycoprotein hormone, chorionic gonadotrophin (CG). These heterodimeric hormones possess a common α-subunit expressed by the CGA gene. Hormone-specific β-subunits, expressed by the FSHB, LHB, TSHB or CGB gene respectively, determine the identity of the heterodimer.

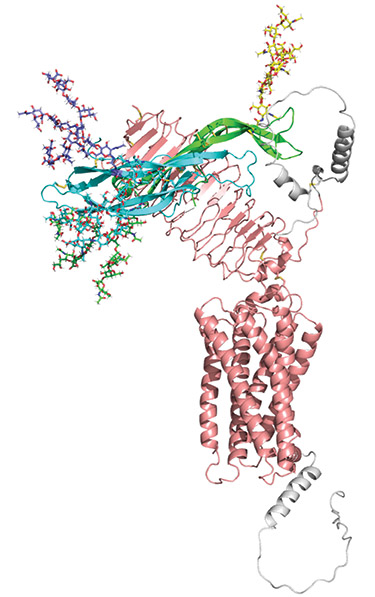
Figure. Fully glycosylated FSH model with the AlphaFold model of FSHR, illustrating relative sizes of FSH glycans and the hormone, and apparent lack of direct glycan contact with receptor. The AlphaFold model was used because some of the FSHR is missing in the structure. FSHα, green; FSHβ, cyan; FSHR, pink; residues missing from cryo-EM structures, grey. Glycan models were created in GLYCAM (www.glycam.org) and are shown as sticks. FSHα: Asn-52, green; Asn-78, yellow. FSHβ: Asn-7, cyan; Asn-24, blue.
Both subunits are glycoproteins decorated with N-glycans at αAsn-52 and αAsn-78 in the α-subunit (the human hormone numbering will be used) and at one or two highly conserved β-subunit Asn residues. The latter differ in numbering due to N-terminal sequence length differences between β-subunits. N-Glycans are highly variable in structure because of a complex biosynthetic pathway that produces differences in glycan branch number and branch termination with negatively charged sialic acid or sulfate residues. Partial N-glycosylation occurs in β-subunits possessing two N-glycosylation sites. Variations in glycan structure and glycan number combine to produce isoforms, of which FSH isoforms have been most extensively studied.
THE IMPACT OF GLYCOSYLATION
The fact that glycoprotein hormone glycosylation is necessary for full activation of Gs by their cognate receptors has been known for over 40 years. While chemically deglycosylated LH receptor binding was not impaired, activation of the Gs-mediated cyclic AMP pathway was compromised.1 Individual subunit deglycosylation revealed that α-subunit glycans exerted a greater impact on biological activity of the heterodimer than β-subunit glycans.1 Site-directed mutagenesis of human CG (hCG) glycosylation sites confirmed the greater influence of α-subunit glycans, localised the critical oligosaccharide to αAsn-52, and revealed a complicated pattern of N-glycan influence on bioactivity.2 Loss of any or all glycans other than that at αAsn-52 had no impact on biological activity. However, only partial loss of activity resulted from αAsn-52 glycan absence. Maximum activity loss required elimination of both αAsn-52 and βAsn-30 glycosylation sites.
Nevertheless, no naturally occurring isoforms exhibited loss of αAsn-52 glycans. Viktor Butnev proposed that the number or size of complex branches in αAsn-52 glycans correlated with biological activity based on studies with equine gonadotrophins: eLH, eFSH and eCG.3 As hCGα, hLHα and hFSHα Asn-52 glycans possess 1, 2 and 3 complex branches respectively, Wendy Walton set out to prepare hFSHα isoforms to test the branch number hypothesis. Along the way, she discovered naturally occurring hFSH variants lacking β-subunit glycans.4
β-SUBUNIT GLYCAN VARIATION
Western blots identify three FSHβ bands at 18, 21 and 24kDa, which represent the lack of Asn-7 glycan, absence of Asn-24 glycan and presence of both glycans respectively. Protein sequencing revealed Asn residues at non-glycosylated sites, indicating oligosaccharyl transferase had skipped these sites. Mass spectrometry confirmed partially and fully glycosylated FSHβ variants and revealed a non-glycosylated FSHβ in human pituitary extracts. Thanks in large part to a FSH glycoform transgenic mouse model developed in Raj Kumar’s laboratory, the three physiologically relevant FSH glycoforms (named after FSHβ sizes) are FSH18, FSH21 and FSH24.5
An age-related switch from predominantly FSH21 to predominantly FSH24 occurs in female pituitaries. Partially and fully glycosylated FSH exhibit divergent patterns of serum concentrations during the menstrual cycle. FSH18 and FSH21 exhibit greater FSHR binding and biological activity than FSH24. The latter engages the receptor more slowly and occupies fewer receptors than preparations containing both FSH18 and FSH21.
Despite support for a trimeric FSHR model based on X-ray crystallography of the FSHR extracellular domain,6 hormone displacement studies suggest most FSHRs exist as monomers.7 PD-PALM studies in Kim Jonas’ laboratory have revealed that 30% of the FSHRs are oligomerised, while FSH binding temporarily reduced this to 20%.8 FSH21/18 mixtures achieve this more rapidly than FSH24. A biased FSH agonist, devoid of cyclic AMP/protein kinase A activity, increases FSHR oligomerisation.
UNDERSTANDING RECEPTOR ACTIVATION
G protein-coupled receptor conformational flexibility challenges structural studies. The glycoprotein
'As cryo-EM is compatible with glycosylated ligands, this technology may eventually provide mechanisms to explain the varying biological activities of glycoprotein hormone isoforms.'
hormone receptors, including the FSHR, are no exceptions. X-ray crystallographic studies were limited to the FSHR extracellular domain (ECD) bound to deglycosylated FSH in order to obtain defractable crystals.9 These studies defined protein–protein interactions involved in high affinity binding to the rigid ECD and interactions with the flexible hinge region.
Recent cryogenic electron microscopic (cryo-EM) reports describe largely complete structures for all three glycoprotein hormone receptors and reveal a common difference between the inactive and active conformations. The inactive receptor ECD is oriented downwards towards the membrane in the inactive state and rotated 38–48° upwards in the active, ligand-bound state.
Two different mechanisms for receptor activation have been proposed. The push–pull model, based on studies of all three receptors, proposes that when the ligand binds, a β-subunit steric clash with the membrane pushes the receptor ECD upwards from the down conformation and receptor hinge interactions with the ligand α-subunit pull the ECD to the fully up, active conformation.10
The glycan-propping model, based on studies of the TSH receptor, proposes that the ECDs flip back and forth between up and down positions.11 ECD position alone is sufficient to activate or inactivate the receptor due to concomitant hinge conformational changes regulating transmembrane domain conformation. TSH binds the up conformation and stabilises it by steric clash of the αAsn-52 glycan with the membrane. The same steric clash precludes glycosylated ligand binding when the receptor is in the down conformation.
Contributions by the β-subunit glycans to receptor binding and hormone activity are not explained by either model. As cryo-EM is compatible with glycosylated ligands, this technology may eventually provide mechanisms to explain the varying biological activities of glycoprotein hormone isoforms.
GEORGE R BOUSFIELD
Department of Biological Sciences, Wichita State University, KS, USA
REFERENCES
- Sairam MR & Bhargavi GN 1985 Science 229 65–67.
- Matzuk MM et al. 1989 Journal of Biological Chemistry 264 2409–2414.
- Butnev VY et al. 1998 Biology of Reproduction 58 458–469.
- Walton WJ et al. 2001 Journal of Clinical Endocrinology & Metabolism 86 3675–3685.
- Wang H et al. 2016 Molecular & Cellular Endocrinology 426 113–124.
- Jiang X et al. 2014 Journal of Biological Chemistry 289 14273–14282.
- Urizar E et al. 2005 EMBO Journal 24 1954–1964.
- Agwuegbo UC et al. 2021 Frontiers in Endocrinology 12 765727.
- Jiang X et al. 2014 Molecular & Cellular Endocrinology 108 7172–7176.
- Duan J et al. 2023 Nature Communications 14 519.
- Faust B et al. 2022 Nature 609 846–853.




{kind=link}