FIRST PUBLISHED IN ISSUE 94 (2009)
The concept of hormones as chemical integrators, flashing through the bloodstream to co-ordinate bodily functions, is well over a century old. However, the molecules from which the first steroid hormones developed are estimated to be over 4,000 million years old, and the endocrine system in which they participate has been evolving ever since the first prokaryotic organism existed.
Our understanding of hormone evolution has leapt forward with the application of genome screening to phylogenetics and the emergence of molecular cladistics. Here, I briefly survey some elementary concepts of hormone evolution from the perspective of a steroid ‘anorak’.
As an unreconstructed steroid biochemist, I reserve the right to insist that steroid hormones rule, OK? Some pundits claim that steroids did not need to evolve – that they are in fact universal bioregulators that came with the big bang. It has been pointed out that steroids are omnipresent throughout the biosphere of the earth and the cyclopentanoperhydrophenanthrene nucleus is the ultimate primordial biomolecule. Some of the most primitive unicellulate organisms have steroids and many bacteria biosynthesise and utilise these and related molecules. Even plants and invertebrates share the familiar acetyl-coA-mevalonate-cholesterol-pregnenolone pathway.
I particularly like the idea that endocrine steroid signalling may have co-evolved with cyclic AMP signalling. Steroids and cyclic AMP are thought to have co-existed in the planetary prokaryotic clone from which the first eukaryotes evolved.
Mycetozoan taxa provide model systems for studying the origin and evolution of eukaryotes. The cellular slime mould (dictyostelid) has a vegetative growth phase during which individual cells exist as amoeba that feed on bacteria. When the food supply is exhausted, they aggregate to form a slug-like plasmodium, which develops into a spore-containing fruiting body. Once food becomes available, the spores are released to produce individual amoeba and the lifecycle starts anew.
The soluble communication factor that prompts amoeba to aggregate into the plasmodium is none other than ‘second messenger’ cyclic AMP. It has been posited that cyclic AMP was a prototypic extracellular bioregulator that allowed elementary cell-to-cell communication but was unable to function as a true hormone owing to its chemical lability. On the other hand, the cell surface membranes of primordial unicellular organisms were probably composed of terpenoid derivatives such as steroids and retinoids that gained increasingly diverse signalling roles as metazoans emerged from the primordial gloop.
Present-day prokaryotes and eukaryotes share common biochemical mechanisms of steroid synthesis, action and catabolism. Thus the basic machinery necessary to evolve tissue-specific cell-to-cell signalling via steroids has been preserved across the plant and animal kingdoms. And throughout evolution, cyclic AMP has retained its quintessential communications function in the endocrine system.

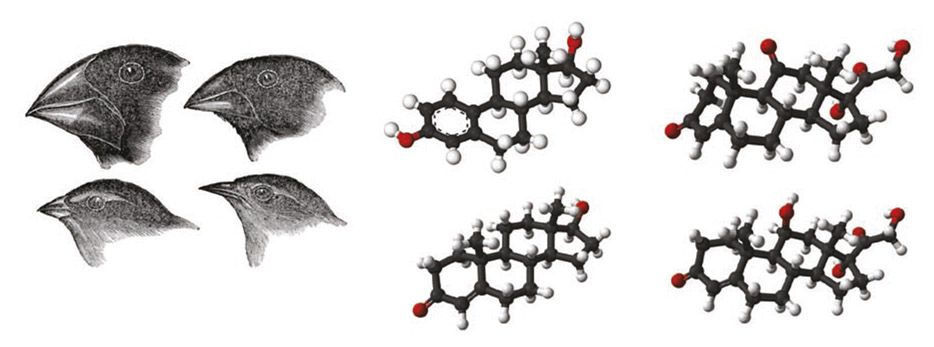
Figure. Darwin noted that finches on the Galápagos Islands had beaks of various sizes and shapes that were highly adapted to different food sources. Subtle differences in the stereochemical structures of steroid hormones such as those illustrated confer radical differences in target-tissue specificity and potency, consistent with an evolutionary pattern of steroid signalling in the endocrine system.
The stereochemical determinants of steroid hormone specificity and potency prompt comparison with the impact of beak morphology on the adaptation of Darwin’s finches to their various food sources on the Galápagos Islands (see Figure).
We now know that paracrine cell-to-cell signalling involving bone morphogenetic protein-4 was responsible for the variation in beak size and shape among the finches he collected. Who knows, had Darwin been a steroid endocrinologist – in the right lab at the right time – correlation of steroid structure and function might have provided him with equivalent insights on natural selection to the Galápagos finches.
Hormone science has evolved beyond recognition since the term hormone was first coined in 1905. And 150 years after On the origin of species, pathway biology has emerged as the ‘fifth force’ in endocrinology, permitting mapping of the cellular receptors, downstream signalling molecules (including cyclic AMP) and attendant changes in gene expression through which hormones are produced and act on their cellular targets. Thus endocrinology has evolved into a diverse and dynamic specialty that integrates the molecules, mechanisms and medicines upon which post-genomic biomedicine is based. Surely, Darwin would have been impressed.
STEPHEN G HILLIER
FURTHER READING
Sandor T & Mehdi AZ 1979. In Hormones and Evolution, vol 1, pp 1–71. London: Academic Press.
Kushiro T et al. 2003. Nature 422 122.
Hillier SG 2005. Journal of Endocrinology 184 3–4.
Abzhanov A et al. 2004. Science 305 1462–1465.



{kind=link}